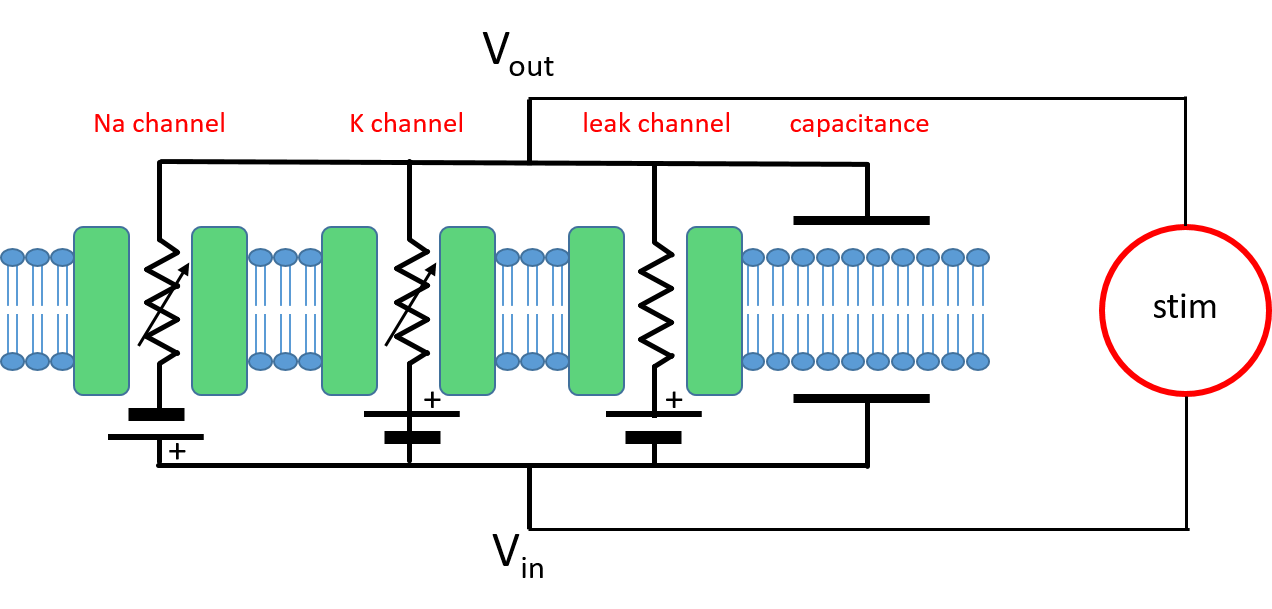
The starting point of the model is that the nerve membrane (specifically, the membrane of the squid giant axon) contains 3 key biological components.
The membrane is thus electronically equivalent to a parallel resistor capacitorSince the model mainly considers the ion channels as conductors, the circuit might better be called a conductor-capacitor circuit. But resistor-capacitor is the term used in electrical engineering, so we will stick with it here. (RC) circuit with additional batteries The model also contains a user-controlled external stimulus acting as a current source. In the real system this could take the form of synaptic input, but the canonical HH model does not include synapses, so some experimental stimulus is necessary to perturb the system and produce a response (otherwise it would just remain in the resting state indefinitely).
In the HH model each voltage-dependent channel can be pictured as a tunnel through the membrane with a small number of gates arranged one-after-another within it. Each gate is either open or shut; there is no intermediate partially-open state, and the transition between states occurs instantaneously. In order for the individual channel to be open and allow ions to flow through, all the gates within that channel must be open simultaneously. If even one gate is shut, then the whole channel is shut. As with the gates, there is no intermediate conductance for the channel; it is either open (full conductance) or shut (zero conductance).

The individual gates flicker between the open and closed state randomly, but the probability of a gate being open is dependent on the voltage across the membrane. In molecular terms, the gates are thought to act like charge-carrying particles, and hence the position they occupy within the membrane, which determines whether they are open or shut, is affected by the electrical potential across the membrane (the voltage).
Channel gates falls into two classes; activation gates which have an open probability that increases with depolarisationDepolarization means that the inside of the cell becomes more positive. The term derives from the fact that at rest the membrane is polarized, with the inside negative relative to the outside. Making the inside more positive is a reduction in that polarization - hence it is depolarized., and inactivation gates which have an open probability that decreases with depolarisation. The probability of a gate being open at any point in time is known as the activation variable for that gate. Since the activation variable defines the probability that a single gate of that class will be open, it therefore also defines the proportion of gates in the total population of that class which are open. As well as differing in how their activation variables change with voltage, gate classes also differ in the rate at which their activation variables change when the voltage changes.
The HH model proposes that each Na channel contains a set of 3 identical, rapidly-responding, activation gates (the m-gates), and a single, slower-responding, inactivation gate (the h-gate ). These two classes of gates in combination explain the transient increase in Na conductivity which results from membrane depolarisation. The way this works is as follows.
At resting potential, the h-gate is open, but the m-gates are shut, and therefore the channel itself is shut (at least, this is the most likely state of affairs; since the gates open and shut probabilistically the state of any gate cannot be predicted with absolute certainty). If the membrane is then depolarised, the m-gates rapidly open, and for a while the channel itself is open or activated.
After a short delay the h-gate shuts, and therefore the channel shuts, even though the membrane is still depolarised. The channel is now in the inactivated state. If the membrane is now repolarised, the m-gates rapidly shut. At this point, if the membrane is again depolarised, the m-gates open, but the h-gate, which has not yet reopened in response to the earlier repolarisation, remains shut, and so the channel itself does not reopen. This is the basis of the absolute refractory period of the action potential. Finally, if the membrane is again repolarised the m-gates shut, and if the membrane is held repolarised for some time, the h-gate eventually reopens (de-inactivation). The channel is now back in its original condition; shut, but ready to open in response to depolarisation.
The K channel is somewhat simpler. It contains a single class of gate consisting of 4 individual activation gates (the n-gates), which respond more slowly than the activation gates of the Na channel. Thus if the membrane is depolarised, the n-gates open, and the K channel opens, but this lags behind the opening of the Na channels,. The K channel has no inactivation gate and so remains open for as long as the membrane remains depolarised. When the membrane is repolarised, the n-gates, and hence the K channel, shut. Again, the response is relatively slow, which means that there is an elevated K conductance for some time following an action potential. This causes an after-hyperpolarisation that is partly responsible for the relative refractory period.
The HH model is one of the most successful mathematical models of a complex biological process that has ever been formulated. The basic concepts expressed in the model have proved a valid approach to the study of bio-electrical activity from the most primitive single-celled organisms such as Paramecium, right through to the neurons within our own brains. Extensions of the basic model, incorporating a variety of voltage-dependent channel types beyond the original HH pair, continue to be very widely used in neuroscience research to this today. However, as HH were themselves well aware, the success of the model does not in itself constitute convincing evidence that the "pictorial" interpretation of the HH equations is a true reflection of the real molecular events.
The main "flaw" in the HH model is the assumption of separate and independent voltage-dependent activation and inactivation gates in the channels. Modern studies on the molecular structure of the channels show that there are indeed charged amino acids within the channel structure that cause a conformational change such that a membrane-spanning channel pore tends to open with a depolarizing voltage change, but neither the Na nor the K channel contain multiple activation gates that operate independently of each other. However, the kinetic properties of the voltage-dependent pore opening and closing closely resemble those that would be produced by multiple independent gates if they existed. In that sense, one might think that HH got lucky. The Na channel does contain an actual inactivation gate (a mobile chain of amino acids) that can block the pore (the ball-and-chain model), but this is not completely independent of activation - the inactivation gate is much more likely to close if the Na channel is already in the activated state, i.e. the pore is open.
Another simplifying assumption in the RC-circuit model is that the channels act like Ohmic (non-rectifying) resistors (reciprocal conductors). In fact, most ion channels show some degree of rectification, but this only becomes significant if there is a very low concentrationIn the extreme case, if the concentration of the permeant ion is 0 on one side of the membrane, that ion clearly cannot flow from the 0 concentration side to the non-0 side! of the permeant ion on one side of the membrane.
The HH model is an exemplar of the aphorism "all models are wrong, but some are useful". The HH model may be wrong in a few details (although right in many more), but it has undoubtedly proved to be extremely useful.